![]() Галерея
Галерея
![]() Галерея
Галерея
Общая характеристика жгутиковых
Обширный класс жгутиковых включает более 6000 видов разнообразных простейших.
Тело жгутиковых имеет весьма разнообразную, чаще овальную, шаровидную или веретеновидную форму (рис. 1). Органеллами движения служат цитонлазматические нитевидные жгутики. Чаще всего их один или два, но может быть четыре и больше. Жгутики обычно помещаются на переднем конце тела и направлены вперед, по движению животного. Однако у некоторых форм при наличии двух и более жгутиков один из них может загибаться вдоль тела назад и иметь значение руля. Этот жгутик обычно соединен с поверхностью тела тончайшей плазматической пленкой, укрепляя таким образом колеблющуюся, или волнообразную, перепонку. Движения направленных вперед жгутиков довольно сложны и разнообразны. Жгутик совершает вращательные движения, описывая фигуру в форме конуса. В то же время он сам волнообразно изгибается. Движение жгутика вызывает вращательное движение тела простейшего, которое как бы ввинчивается в воду. У некоторых жгутиковых (отряд Rhizomastigida), кроме постоянных жгутиков, имеются псевдоподии. Эти формы указывают на близость классов жгутиковых и саркодовых.
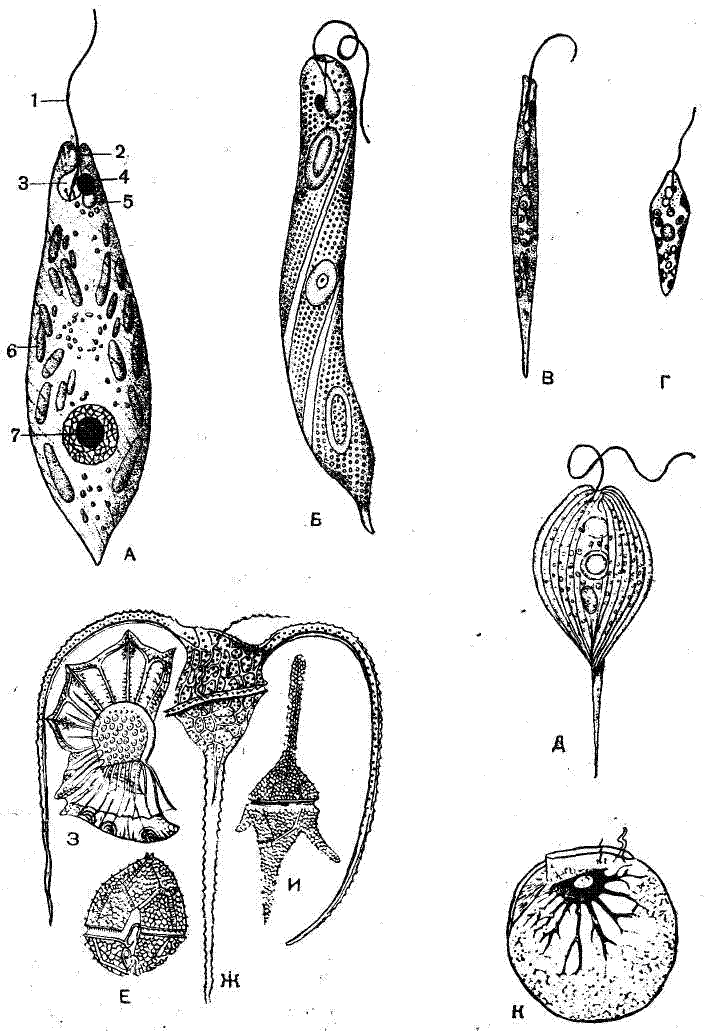
Рис. 1. Различные жгутиковые: |
Тело жгутиковых состоит из цитоплазмы и обычно одного пузыревидного ядра (рис. 1). Цитоплазма разделяется на два слоя: наружный, более плотный, гомогенный и прозрачный — эктоплазму и внутренний, более жидкий и зернистый — эндоплазму.
Тело большинства жгутиковых покрыто довольно плотной эластичной оболочкой — пелликулой, образующейся за счет уплотнения эктоплазмы. Пелликула позволяет животному временно изменять форму тела, или метаболировать. У некоторых жгутиковых тело покрыто твердой оболочкой из клетчатки, которая образует своеобразный панцирь.
Типы питания простейших наиболее разнообразны в классе жгутиковых. Только среди них встречаются формы, способные, подобно растениям, к фотосинтезу, т. е. к синтезу различных углеводов из ранее поглощенных углекислого газа и воды, происходящему на солнечном свету. Белки их тела строятся из синтезированных углеводов и поглощенных из окружающей среды неорганических солей, в том числе азотистых. Таких жгутиконосцев называют аутотрофными, а их способ питания голофитным или растительным.
К жгутиковым с голофитным типом питания относятся все организмы, имеющие хлорофилл. Они питаются так же, как и все зеленые растения. Органеллами питания у таких жгутиковых служат окрашенные тельца — хроматофоры различной формы и величины (рис. 1, А), содержащие хлорофилл и другие пигменты (желтый, красный). Благодаря маскирующему влиянию этих пигментов многие жгутиковые не имеют зеленой окраски, несмотря на наличие хлорофилла. Жгутиковые с голофитным питанием часто имеют уже упомянутую выше клетчатковую оболочку. Это еще более подчеркивает их принадлежность к растениям.
Другие жгутиковые питаются органическими продуктами распада более сложных веществ, которые осматическим путем поступают внутрь их тела. Такой тип питания называют сапрофитным, а организмы, питающиеся таким способом,— сапрофитами. Сапрофиты уже не аутотрофные (самопитающиеся) организмы, а гетеротрофные, т. е. питающиеся готовым органическим веществом. Жгутиковые сапрофиты живут в загрязненых водоемах, богатых разлагающимися органическими веществами. Потребляя их, жгутикбвые тем самым участвуют в очистке воды подобных водоемов.
Многие паразитические жгутиковые и другие простейшие питаются также сапрофитным способом, всасывая всей поверхностью тела сыворотку крови, тканевые жидкости или (кишечные паразиты) продукты переваривания пищи их хозяина.
Наконец, третью группу составляют жгутиковые, питающиеся, как типичные животные, твердой пищей: бактериями, мелкими водорослями и т. п. Они имеют специальные органеллы питания: рот, глотку и пищеварительные вакуоли; последние образуются в плазме по мере заглатывания пищи. Такой тип питания называют голозойным или анимальным.
Особое положение по способу питания занимают некоторые эвглены, в частности зеленая эвглена (Euglena viridis) (рис. 1, А). У зеленой эвглены имеются хроматофоры, содержащие хлорофилл. На свету она питается как типичный зеленый растительный организм. Но, помещенная в темноту, эвглена обесцвечивается и переходит к сапрофитному питанию. По-видимому, и на свету эвглена обладает смешанным способом питания, т. е. является миксотрофным организмом.
Голофитные жгутиковые обычно имеют красное или бурое пятнышко— глазок, или стигму,— расположенное на переднем конце тела у основания жгутика (рис. 1, А). Это — светочувствительная органелла, которая обусловливает способность зеленых жгутиковых воспринимать световые раздражения. Зеленые жгутиковые обнаруживают положительную реакцию на свет, т. е. обычно передвигаются по направлению к источнику света.
У многих пресноводных жгутиковых имеются специальные органеллы выделения — сократительные вакуоли, при помощи которых из клетки удаляется избыток воды, а также жидкие продукты диссимиляции. Сократительная вакуоля у эвглены (рис. 1, А) помещается в передней части тела. Сократительная вакуоля периодически наполняется жидкпм содержимым, которое при сжатии вакуоли выталкивается в особый пузырек — резервуар (рис. 1, А), а затем через особый канал, иногда неправильно называемый «глоткой», выделяется наружу. Сократительные вакуоли, как правило, отсутствуют у морских жгутиковых. Кроме описанных органелл, в цитоплазме жгутиковых могут быть различные включения: зерна крахмала (у зеленых жгутиковых), зерна парамила — углевода, близкого по составу к крахмалу (у эвглены и др.), капельки жира и т. п.
Для большинства жгутиковых известно только бесполое размножение. Половой процесс изучен лишь у зеленых колониальных форм из семейства вольвоксовых (отряд Phytomonadida).
Бесполое размножение жгутиковых происходит обычно путем деления надвое. Оно начинается с митотического деления ядра, за которым следует продольное деление тела, начинающееся с его переднего конца (рис. 2). При этом делятся также базальные тельца; жгутики или отбрасываются, или переходят к одной из дочерних особей, а у другой развиваются заново из базального тельца. При делении жгутиковых, имеющих хроматофоры, последние также делятся и распределяются по дочерним клеткам. У некоторых жгутиковых делению предшествует инцистирование, при этом деление происходит уже под оболочкой цисты (рис. 2, 2 и 3).
 Рис. 2. Размножение зеленой эвглены: 1 — продольное деление; 2 и 3 — дедеаие в цисте. |
У колониальных жгутиковых из вольвоксовых при бесполом размножении
отдельная клетка колонии делится несколько раз (по числу клеток составляющих
колонию), и таким образом возникают дочерние колонии.
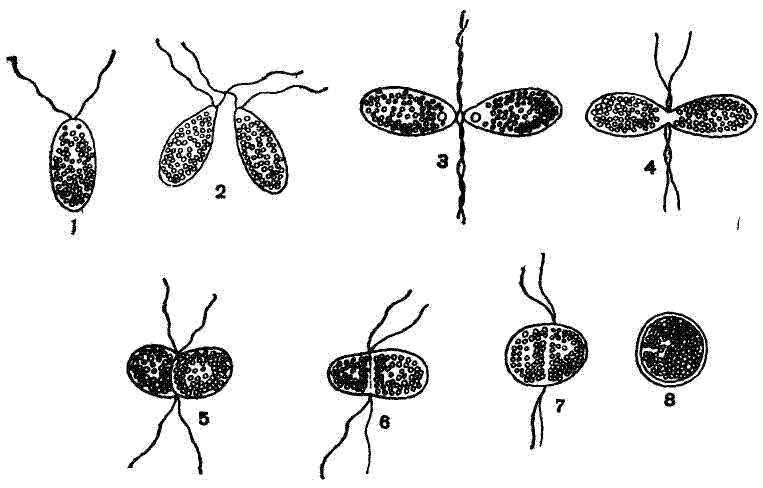 Рис. 3. Изогамная копуляция у Polytoma uvella: 1— гамета, 2—7 — последовательные стадии копуляции гамет, 8 — инцистированная зигота. |
Половое размножение у разных жгутиковых происходит различно. В простейшем случае происходит копуляция двух внешне одинаковых по величине половых особей — гамет, например у Polytoma (рис. 3 ) и некоторых видов хламидомонад. Такую копуляцию одинаковый гамет, или изогамет, называют изогамной копуляцией. У других жгутиковых гаметы отличаются размерами. Одни из них более крупные — макрогаметы, другие более мелкие — микрогаметы (рис. 4). Макро- и микрогаметы снабжены жгутиками. Макро- и микрогаметы копулируют друг с другом. Копуляцию неравных гамет, или анизогамет называют анизогамной.
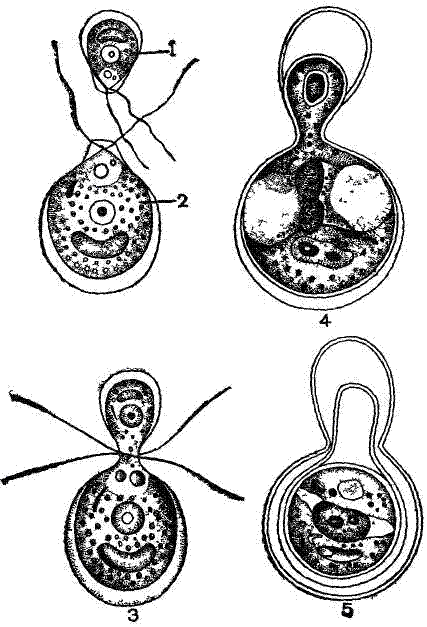 Рис. 4. Анизогамная копуляция у Chlamydomonas braunii: 1 — микрогамета, 2 — макрогамета, 3—5— последовательные стадии копуляции гамет. |
Формирование анизогамет можно считать первым этапом дифференциаций пола. У колоний Volvox, образованных огромным числом особей (до 10—20 тыс.), гаметы могут формироваться только из отдельных генеративных особей. Большинство особей колонии вегетативные, не способные участвовать в половом размножении. В колонии Volvox только 25—30 клеток (особей) может превращаться в крупные неподвижные макрогаметы и лишь 5—10 клеток—образовывать микрогаметы (рис. 5). При формировании макрогамет деления клеток не происходит, а при образовании микрогамет происходит ряд делений, в результате которых возникают 256 мельчайших микрогамет, каждая с парой жгутиков. Микрогаметы покидают колонию и выходят в воду. Затем они проникают в другие колонии, где копулируют с макрогаметами.
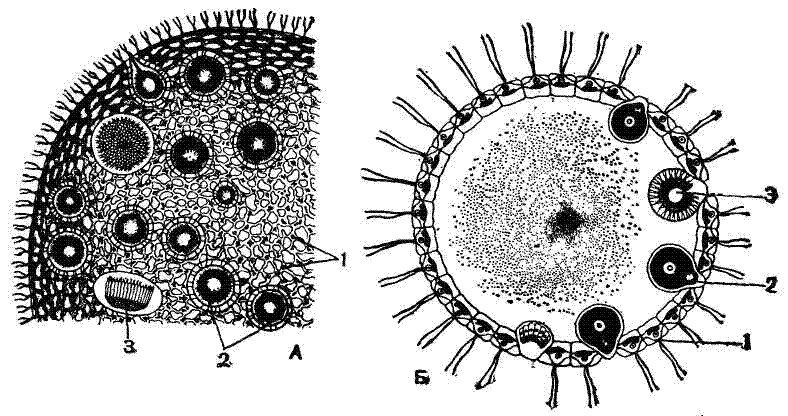
Рис. 5. Вольвокс (Volvox globator): |
Макрогаметы жгутиками не обладают. Их можно сравнить с яйцевыми клетками многоклеточных, а микрогаметы — со сперматозоидами. Такую копуляцию Volvox называют оогамией. После оогамнои копуляции образуется оплодотворенная макрогамета— зигота. Последняя выделяет плотную оболочку, а все вегетативные особи колонии погибают. Затем зигота начинает многократно делиться и дает начало новой колонии Volvox.
У Volvox globator микро- и макрогаметы развиваются в одной и той же гермафродитной колонии, тогда как у другого вида — V. aureus — колонии раздельнополы: одни из них (женские) образуют только макрогаметы, другие (мужские) — только микрогаметьь.
Таким образом, у различных представителей жгутиковых наблюдаются как бы различные ступени эволюции полового процесса — от изогамной копуляции до настоящей оогамии, что представляет интерес для понимания происхождения полового процесса у многоклеточных.
Класс жгутиковых (Mastigophora, или Flagellata) состоит из большого числа (13—14) отрядов. Главнейшие из них следующие: 1. Эвгленовые (Euglenida); 2. Панцирные (Dinoflagellata); 3. Фитомонадовые (Phy-tomonadida); 4. Корнежгутнковые (Rhizomastigida); 5) Протомонадовьш (Protomonadida); 6. Многожгутиковые (Polymastigida); 7. Гипермастигиновые (Hypermastigida); 8. Опалиновые (Opalinida).
Первые три отряда (эвгленовые, панцирные, фитомонадовые) входят в состав подкласса фитомастигины (Phytomastigina) — аутотрофных растительных жгутиконосцев. Следующие четыре отряда (корнежгутиковые, протомонадовые, многожгутиковые, гипермастигиновые) составляют подкласс зоомастигины (Zoomastigina) — типичных гетеротрофных организмов. Последний отряд (опалиновые) многие ученые выделяют в ранг самостоятельного подкласса Opalinina.
Литература: Зоология беспозвоночных. Учебник для студентов биол. фак. пед. ин-тов / Натали В. Ф. - издательство "Просвещение" 1975 год
![]() Счетчик
Счетчик
Жгутиковые (Mastigophora, или Flagellata)
![]() Наши партнеры
Наши партнеры
![]() Поиск
Поиск
![]() Меню
Меню
| Пн | Вт | Ср | Чт | Пт | Сб | Вс |
| 1 | 2 | 3 | 4 | 5 | 6 | |
| 7 | 8 | 9 | 10 | 11 | 12 | 13 |
| 14 | 15 | 16 | 17 | 18 | 19 | 20 |
| 21 | 22 | 23 | 24 | 25 | 26 | 27 |
| 28 | 29 | 30 | 31 | |||
![]() Календарь
Календарь