![]() Галерея
Галерея
![]() Галерея
Галерея
Автор: Гайнанов Константин
Сайт: BioMirOk.narod.ru
Кожа, будучи неповрежденной, непроницаема для большинства инфекционных агентов. Слизь, выделяемая стенками внутренних органов, действует как защитный барьер, препятствующий прикреплению бактерий к эпителиальным клеткам. К другим механическим факторам, способствующим защите поверхности эпителия, можно отнести вымывающее действие слез, слюны и мочи. Совершенно иной характер имеет механизм микробного антагонизма, связанный с нормальной бактериальной флорой человека. Эти бактерии угнетают рост многих патогенных микроорганизмов и грибов вследствие конкуренции. Если же инфицирование организма все же произошло, то в действие вступают иммунологические способы защиты:
1. Гуморальные: разрушение растворимыми химическими факторами и
Эти иммунологические способы защиты в совокупности и составляют основу иммунологии. Иммунную систему поэтому можно рассматривать как совокупность систем, обеспечивающих эти способы защиты. Соответственно, иммунная система состоит из лимфоцитов (B- и T-лимфоцитов), макрофагов и ряда сходных с макрофагами клеток. Клеточные элементы иммунной системы организованы в тканевые и органные структуры, к которым относятся селезенка, лимфатические узлы, пейеровы бляшки кишечника, миндалины, тимус и костный мозг. Кроме того весьма значительная часть лимфоцитов и макрофагов составляет популяцию клеток крови и лимфы. Стимуляция клеточного иммунитета с неизбежностью приводит к снижению эффективности гуморальных факторов защиты и наоборот.
Реакции, которые непосредственно вызывают разрушение патогенных факторов, составляют в совокупности эффекторные механизмы иммунной системы .
Возбудители бактериальных или вирусных заболеваний в процессе своей жизнедеятельности в организме хозяина оказываются в той или иной ситуации во внеклеточной среде. Пребывание в жидкостях организма может быть более длительным в случае с внеклеточными патогенами или более коротким - при поражении хозяина внутриклеточными бактериями или вирусами, но оно обязательно представлено. При нормальном функционировании иммунной системы патогены и их токсины, оказавшиеся вне клеток хозяина, подвергаются действию антител - эффекторных молекул, продуцируемых В-лимфоцитами. Вопросы, относящиеся к гуморальному иммунному ответу, связаны с функционированием антител и формированием клеточных и молекулярных механизмов эффекторной фазы работы В-системы иммунитета. Участие антител в иммунном ответа проявляется в трех формах:
- активации системы комплемента.
Итак, вирусы и внутриклеточные бактерии для своего воспроизведения должны первоначально проникнуть из жидкостей организма в клетку - место своей жизнедеятельности. Оказавшись, даже короткое время, во внеклеточном пространстве, патогены подвергаются нейтрализующему действию антител. Эта активность антител проявляется в блокаде рецепторного взаимодействия между патогеном и инфицируемой клеткой. Иначе, антитела препятствуют предетерминированному взаимодействию клеточных рецепторов с лигандом на поверхности патогена. Процесс нейтрализации проявляется не только в случаях с корпускулярными антигенами, но и с бактериальными токсинами. Одним из ведущих механизмов элиминации (лат. eliminare - изгонять) внеклеточных патогенов является активность фагоцитирующих клеток, которые, захватывая антиген, разрушают его в фаголизосомах. Этот неспецифический по основной своей сути процесс усиливается специфическими антителами. Макрофаги - главные участники внутриклеточного разрушения патогенов - имеют на своей поверхности рецептор к Fc -фрагменту иммуноглобулинов. Патоген, связавшийся со специфическим антителом, оказывается значительно более доступным для фагоцитирующих мононуклеаров в результате взаимодействия Fc-фрагмента иммуноглобулина с Fc-рецептором на поверхности фагоцита.
Процесс усиления фагоцитоза за счет гуморальных факторов вообще и специфических антител в частности получил название опсонизации. Третья форма функционального проявления антител связана с системой комплемента. Антитела, связавшиеся с поверхностью бактериальной клетки, активируют белки системы комплемента, которые принимают участие в ряде иммунологических явлений:
- во-первых, взаимодейстуя с патогеном, некоторые белки системы комплемента выполняют функцию опсонинов;
- во-вторых, компоненты комплемента выступают в качестве хемотаксических факторов, привлекая в очаг инфекции фагоцитирующие клетки;
- третье свойство белков системы комплемента связано с их литической активностью - способностью образовывать поры в клеточной стенке бактерий, что приводит к гибели патогенов.
При отсутствии антигенной агрессии специфические антитела не образуются. По этому признаку процесс антителогенеза классифицируется как индуцибельное явление. Фактором индукции выступает антиген.
Однако, как и в случае с Т-клетками, одного специфического сигнала от антигена недостаточно для начала синтеза антител В-клетками. Необходим второй сигнал для реализации информации от специфического индуктора. Роль второго сигнала выполняют цитокины , продуцируемые хелперными CD4 T-клетками . Получение второго сигнала для полноценного развития В-клеток в антителопродуцирующие плазмоциты возможно при непосредственном контактном Т-В-взаимодействиии.
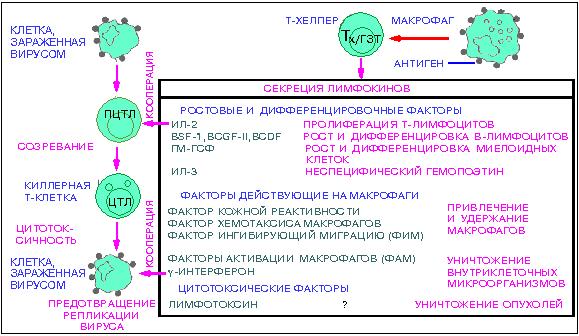
Неспецифический клеточный иммунитет обусловлен главным образом наличием в крови лейкоцитов и их фагоцитарной активностью. Способностью к фагоцитозу обладают гранулоциты, моноциты, тромбоциты и лимфоциты. Сильнее всего фагоцитарная активность выражена у моноцитов, которые содержат большое количество лизосомных ферментов, расщепляющих захваченные частицы. В основе клеточного иммунитета лежит функционирование нескольких субпопуляций T-лимфоцитов, каждый из которых может иметь ту же антигенную специфичность, хотя функции каждой из этих субпопуляций различны. Это аналогично различным функциям разных классов иммуноглобулинов, которые имеют идентичную специфичность, но различные биологические функции. В систему функционирования этих эффекторных T-клеток (T-эффекторов ) входит:
1. Продуцирование T-хелперами разнообразных растворимых факторов (лимфокинов), стимулирующих B-клеточный ответ.
2. Воспалительный эффект. Лимфокины, продуцированные T-хелперами, индуцируют миграцию и активацию моноцитов и макрофагов, что вызывает острую воспалительную реакцию .
3. Пролиферация цитотоксических T-лимфоцитов. Цитотоксические T-лимфоциты сами по себе способны уничтожать другие клетки, прежде всего клетки, зараженные вирусом. При этом рецепторы цитотоксических T-лимфоцитов распознают поверхностные антигены на клетках-мишенях, но меньшей степени, чем антитела , способные распознавать разные штаммы вирусов, т.е. обладают меньшей специфичностью.
4. Если же некоторой части вирусов все же удается уйти из под контроля цитотоксических T-лимфоцитов в виде "раздетой" вирусной частицы в соседние клетки, то система клеточного иммунитета использует другой механизм их инактивации: T-хелперы, стимулированные вирусным антигеном, выделяют лимфокин гамма-интерферон , который придает соседним клеткам непермиссивность для репликации любого проникшего в них вируса. Гамма-интерферон может, кроме того, повышать неспецифическую цитотоксичность NK для зараженных клеток.
5. Регуляторные эффекты. T-супрессоры могут индуцировать супрессию иммунного ответа других эффекторных клеток.
 |
Изучение генетического контроля силы иммунного ответа как самостоятельного направления исследований началось в 60-е годы и вскоре слилось с проблемой, направленной на выяснение механизмов распознавания антигена Т-клетками. Первые опыты по изучению генетической детерминации силы иммунного ответа были проведены с беспородными кроликами в первой трети нашего столетия. Исследования установили наследуемость иммунного ответа по доминантному типу, однако они не получили развития. Для полноценного изучения явления необходимо соблюдение, по крайней мере, двух условий:
- наличие генетически хорошо охарактеризованных животных, какими являются, в частности, инбредные линии мышей и морских свинок, и
- контролируемые по специфичности антигены.
Тем не менее было уже известно, что разные индивиды неодинаково реагируют на один и тот же антиген. Результаты семейного анализа чувствительности с Corynebacterium diphtheriae позволили показать, что резистентность или чувствительность к инфекции может быть наследуемым признаком. В пользу этого свидетельствовали и данные, полученные на морских свинках разных линий: Фьерд-Шайбель (1943г.) продемонстрировал, что продкуцию дифтерийного антитоксина контролирует один ген, и она наследуется как доминантный менделевский признак. В этом же эксперименте был установлен также доминантный тип наследования высокой иммунореактивности.
К началу 60-х годов в распоряжении экспериментаторов было уже достаточное количество инбредных, конгенных и рекомбинантных линий мышей, а также целый набор искусственно синтезированных пептидов с ограниченной специфичностью вплоть до моноспецифичности. Это позволило подойти к решению проблемы генетического контроля силы иммунного ответа и изучению четырех основных вопросов его регуляции:
1) каков характер наследования силы иммунного ответа,
2) где в геноме клетки локализованы гены иммунного ответа (Ir-гены),
3) что представляют из себя фенотипические продукты этих генов,
4) в каком типе клеток экспрессируются продукты, контролируемые Ir-генами.
Литература:
1) Васильева В.И., Волков И.М., Ярыгин В.Н.,
Синельщикова В.В. Биология. 2 кн. Кн. 1. М: Высшая школа, 2004год – 431с.
2) Лобашев М.Е. Генетика. Изд. Ленинградского
университета, 1969год – 750с.
3) Мамонтов С.Г., Захаров В.Б., Сивоглазов В.И.
Биология. Общие закономерности. М.: «школа – пресс», 1996год – 624с.
4) Мюнтцинг А. Генетика. Изд. Мир, Москва, 1967 –
610с.
5) Солодова Е.А., Богданова Т.Л. Биология.
Справочник. – М.: АСТ – ПРЕСС ШКОЛА, 2002 – 816с.
![]() Счетчик
Счетчик
Клеточная и гуморальная система иммунитета, генетический контроль
![]() Наши партнеры
Наши партнеры
![]() Поиск
Поиск
![]() Меню
Меню
![]() Календарь
Календарь